МІКРОХИМЕРИЗМ. I. МОЖЛИВІ ДЖЕРЕЛА Й РОЛЬ ПРИ АУТОІМУННИХ ЗАХВОРЮВАННЯХ
Гринь В.К., Гнилорыбов A.М., Иващенко Е.С. , Трубникова Н.Н. , Мелёхина B.А., Полулях О.Е., Риджок В.В.
Резюме. У статті обговорюється нове у сучасній медицині та ревматології явище — мікрохи-меризм. Наведено історію вивчення мікрохиме-ризму, його можливі джерела, варіанти (фетальний, материнський тощо), роль мікрохимеризму у розвитку деяких аутоімунних і ревматичних захворювань, механізми впливу на перебіг патологічних аутоімунних процесів. Окремо представлені останні дані щодо виявлення мікрохимер
та їх клітинний склад при деяких захворюваннях (системна склеродермія, вовчак, дерматоміозит, діабет І типу, злоякісні новоутворення). Показано, що важливе значення у розвитку ревматичних хвороб може належати «близькості» HLA-фенотипів хазяїна і донора мікрохимерних клітин, що призводить до розвитку реакції, що нагадує хронічну реакцію «трансплантат проти хазяїна». Можливе значення приналежності мі-крохимерних клітин до певних типів (наприклад до Т-лімфоцитів). Автори обговорюють можливість терапевтичного використання химеризму для індукції імунологічної толерантності та лікування раку.
ИСТОРИЯ ВОПРОСА И ОПРЕДЕЛЕНИЕ ПОНЯТИЯ
Ничто не совершенно, и иммунная система человека — не исключение. Механизмы иммунитета, поддерживающие генетическую неприкосновенность и однородность, в некоторых ситуациях могут вызывать толерантность, а при других условиях — приводить к системным аутоиммунным реакциям.
Согласно греческой мифологии химера — легендарное существо, считавшееся почти непобедимым, тело которого состояло из частей многих животных (рис. 1). Химера обладала не только своей собственной силой, но и силой других животных, из частей которых она состояла. В наши дни термин «химеризм» прочно вошел в медицинскую терминологию. Этот термин употребляется в тех случаях, когда экспериментальные животные или человек приобретают чужую генетическую составляющую (например в результате трансплантации) и становятся ее постоянным «толерантным» хозяином. При химеризме (например в результате трансплантации органов) количество донорских клеток велико. В период беременности, при трансфузии крови, количество клеток очень незначительно, и это называется микрохимеризмом. Термин «микрохимеризм» означает наличие очень небольшого количества генетически чужеродных клеток в организме хозяина и в последние годы применяется все шире. «Микрохимерные» клетки не только живут в организме хозяина, но и способны к дифференцировке и персистенции. Эти клетки проявляют иммунологическую компетентность и могут дифференцироваться в разных направлениях, что может играть важную роль при аутоиммунных расстройствах и отторжении трансплантата (реакция «трансплантат против хозяина» — РТПХ), раке и некоторых типах репарации тканей.

Рис. 1. Химера Агено (мифическое трехглавое чудовище) — бронзовая фигура, обнаруженная в городе Ареццо (Италия) в 1553 г. (музей археологии во Флоренции)
История микрохимеризма началась с идеи о взаимосвязи перенесенных беременностей и развития аутоиммунных болезней, поскольку последние значительно чаще возникают у женщин в возрасте старше 30 лет — когда большинство из них имеют детей. Известно, что вынашивание детей приводит к развитию микрохимеризма. Кроме того, некоторые аутоиммунные заболевания по своим проявлениям отдаленно напоминают хроническую реакцию «трансплантат против хозяина» (хРТПХ).
Факт фетоплацентарного движения клеток впервые стал известен более 10 лет назад. В 1996 г. доктор Diana Bianchi и соавторы [6] впервые выявили феномен циркуляции фетальных клеток в организме матери через десятилетия после беременности. Микрохимерные клетки были выявлены в щитовидной железе, сердце, периферической крови. В результате распространения значительное количество микрохимерных клеток «заселяет» лимфоузлы.
Но только в последнее время было выяснено, разрушаются ли эти клетки или остаются сохранными и способны ли они далее дифференцироваться. Часть таких клеток действительно выживает, и, предположительно, количество выживших клеток прямо пропорционально совместимости плода и матери.
Первыми исследователями, предложившими идею о роли микрохимеризма в патогенезе аутоиммунных болезней, были J.L. Nelson и соавторы [5]. После эпидемиологических исследований аутоиммунной патологии у женщин (а потом и у мужчин) и в наблюдениях после трансплантаций они выдвинули гипотезу о том, что химерные клетки могут быть причиной многих иммунопатологических феноменов при склеродермии, волчанке, сахарном диабете I типа и др. Авторы описали также «мужской» микрохимеризм у матерей, не имевших в анамнезе беременностей плодом мужского пола. Исходя из этого, авторы сделали вывод о том, что микрохимеризм может быть вызван не только беременностью, но и другими причинами (например передаваться из поколения в поколение, от плода предыдущей беременности или от близнеца).
ИСТОЧНИКИ МИКРОХИМЕРИЗМА
Предполагается существование нескольких возможных вариантов микрохимеризма: феталь-ный (миграция клеток плода в организм матери), материнский (миграция материнских клеток в организм плода), микрохимеризм близнецов (обмен клетками между близнецами), переход из кровотока матери в организм плода клеток от предыдущих беременностей, обмен клетками между супругами, трансплантационный микрохимеризм (как правило, в результате гемотрансфузий).
Некоторые считают, что беременность приводит к иммуносупрессии с развитием толерантности к клеткам плода, то есть к микрохимериз-му (рис. 2). Переход аллогенных фетальных клеток в материнский кровоток называется фетальным микрохимеризмом. Выявление фетальных клеток в материнских тканях на протяжении продолжительного времени после беременности регистрируется значительно чаще, чем миграция материнских клеток в кровоток плода (материнский микрохимеризм) [1, 2]. При нормальном течении беременности фетальный микрохимеризм впервые определяется на 4-6-й неделе гестации [6, 7], а количество фетальных клеток и фетальной ДНК в кровотоке матери увеличивается со сроком ге-стации [3, 4, 8] и особенно во время родов. Фе-тальные клетки могут персистировать затем в организме женщины на протяжении десятилетий после завершения беременности [9].
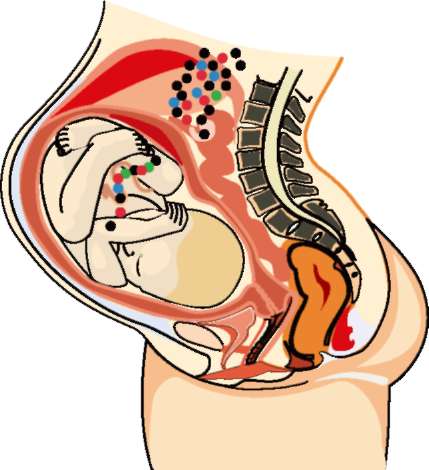
Рис. 2. Схематическая диаграмма беременной с передачей фетальных клеток (черных) матери и материнских клеток (красных) плоду. Материнские стволовые клетки — зеленые, стволовые клетки плода — синие
И кровь плода, и плацента содержат стволовые клетки и клетки-предшественники. Плацента является богатым источником фетальных гематопоэти-ческих стволовых клеток. Плацентарные кровотечения, естественные или ятрогенные повреждения ворсинок трофобласта являются важными причинами клеточного обмена плода и матери [2].
Альтернативный источник микрохимерных клеток — обмен клетками между близнецами, которые также персистируют во взрослой жизни [10]. Впервые микрохимеризм был описан у близнецов в 1957 и 1959 г. [12, 13]. Другой источник — переливание крови. Длительное персистирование донорских клеток определяется у пациентов с множественными гемотрансфузиями после травм [11].
Гипотетически микрохимерные клетки должны быть недифференцированными, поскольку способны циркулировать в организме и заселять различные органы, экспрессировать антигены, характерные для соответствующих тканей. Вследствие этого предполагается, что микрохимерные клетки являются стволовыми [14]. Их назвали ассоциированными с беременностью клетками-предшественниками [15].
Трансплацентарная миграция фетальных клеток цитотрофобласта естественна для нормального развития плаценты и плода, и их (фетальных клеток) наличие в период беременности выполняет важную функцию в индукции иммунологической энергии и/или толерантности у плода и матери [16]. Фетальные и плацентарные аномалии, такие как фетальная анеуплодия, преэклампсия и прерывание беременности приводят к увеличению количества фетальных клеток разных типов в кровотоке матери, и это может повышать вероятность их постоянного присутствия в тканях женщины [17-19].
Известно, что в период беременности материнская кровь содержит разные типы фетальных клеток, включая трофобласты, эритробласты с ядром, тромбоциты, В- и Т-лимфоциты, моноциты, естественные киллеры и некоторые типы клеток-предшественников/стволовых клеток [2, 20-25].
Фетальные мезенхимальные стволовые клетки могут дифференцироваться в остеогенном, хондро-генном, миогенном и адипогенном направлениях. Они определяются в низких концентрациях в I триместр в печени, костном мозгу [22], а также в крови матери после беременности [20]. Кроме этого, фетальные химерные клетки экспрессируют маркеры эндотелиальной дифференцировки (VEGFR2+, CD31+), что наталкивает на мысль об их участии в материнском неоангиогенезе в период беременности [26]. Таким образом, фетальные химерные стволовые клетки/клетки-предшественники могут принимать активное участие в восстановлении поврежденных тканей. С эволюционной точки зрения это явление может быть интерпретировано как помощь плода в защите благополучия матери в период и после беременности. Патологическое значение микрохимерных клеток состоит в их возможном участии в инициировании и/или утяжелении течения аутоиммунных поражений разнообразных органов и тканей (рис. 3).
РОЛЬ МИКРОХИМЕРИЗМА И HLA В РАЗВИТИИ АУТОИММУННЫХ И РЕВМАТИЧЕСКИХ БОЛЕЗНЕЙ
Система HLA (human leukocyte antigen) — неотъемлемая часть иммунного ответа и ключевой момент для успеха трансплантации — занимает центральное место в распознавании организмом «своего» и «чужого». Из-за значительной клинической схожести хР^Х и системной склеродермии, исследования взаимоотношения микрохимеризма и HLA клеток хозяина и чужеродных клеток впервые были проведены при этом заболевании. Первое исследование, описывающее микрохимеризм при этом заболевании, включало количественную оценку фетального микрохимеризма у женщин, оценку HLA у матерей и их детей [27].
Если HLA плода значительно отличается от HLA матери, эти клетки распознаются как чужеродные и разрушаются иммунной системой. Однако, если HLA не идентичны, но достаточно совместимы, они заселяют материнский организм. В таких ситуациях они могут вызывать реакции по типу хР^Х. В связи с этим, согласно одной из первых гипотез, взаимоотношения чужеродных HLA и HLA хозяина играют большую роль в определении последующего патогенного потенциала персистирующих клеток [2В]. В большинстве случаев гены HLA ребенка, унаследованные от отца, отличаются от материнских HLA, таким образом, они оказываются несовместимыми. Иногда ген HLA, унаследованный от отца, совпадает с геном HLA матери, что привело к предположению о повышенном риске заболевания в случае сходства (но не полной идентичности!) HLA матери и ребенка. Повышенный риск заболевания системной склеродермией, ассоциированный с HLA-DRB1-совместимостью, представляет большой интерес в свете того, что реципиенты после гемо-трансфузий от HLA-совместимых членов семьи находились в группе повышенного риска возникновения фатальной P^X [29].
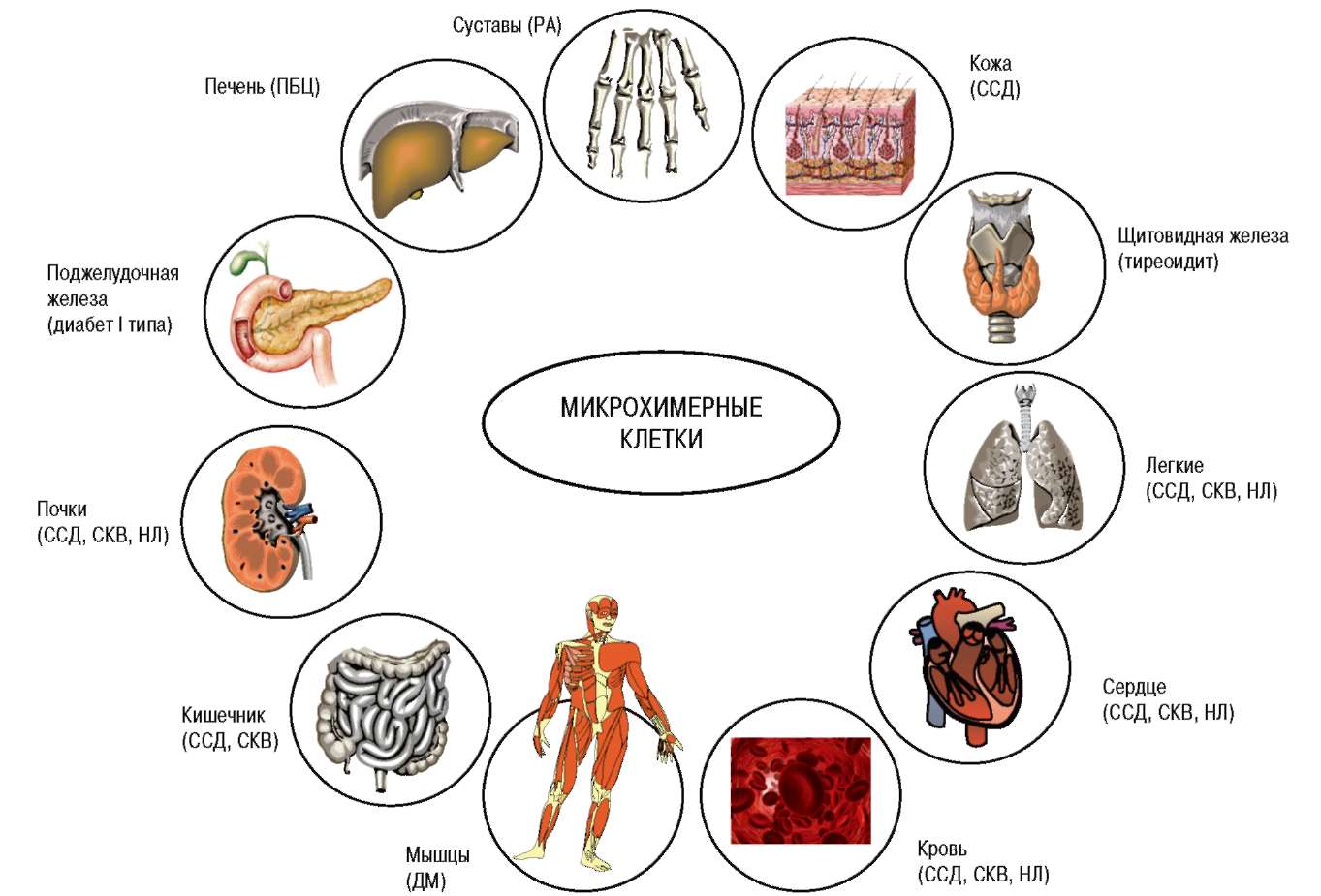
Рис. 3. Органы, в которых присутствие микрохимерных клеток сопровождалось аутоиммунной реакцией. ССД — системная склеродермия; СКВ — системная красная волчанка; ДМ — дерматомиозит; РА — ревматоидный артрит; ПБЦ — первичный билиарный цирроз; НЛ — неона-тальный люпус-синдром
МЕХАНИЗМЫ ВЛИЯНИЯ МИКРОХИМЕРИЗМА НА ТЕЧЕНИЕ БОЛЕЗНЕЙ
Точные механизмы, с помощью которых микро-химеризм может влиять на патогенез аутоиммунных заболеваний, пока не известны. P.J. Christner и соавторы [З0] описали модель, которая подтверждает непосредственную роль микрохимеризма в патогенезе системной склеродермии. В результате тщательных экспериментов исследователи смогли сопоставить микрохимеризм с патологией и с раздражающим внешним фактором (винилхлоридом). Предположительно, хРТПХ обусловлена донорскими Т-клетками, которые не уничтожаются иммунной системой организма хозяина (состояние толерантентности) [31].
Однако имеется существенная разница между xPTПX и микрохимеризмом, развившимся после беременности – это различие состоит в количестве чужеродных клеток в организме хозяина. При системной склеродермии соотношение чужеродных микрохимерных клеток и клеток хозяина составляет примерно 1 на млн лейкоцитов хозяина (или 1 чужеродная клетка на 600 тыс. лимфоцитов хозяина) [27, 32]. В то же время, при xPTПX донорские клетки практически полностью замещают циркулирующие клетки хозяина.
Презентация чужеродных пептидов клетками хозяина друг другу называется «непрямым» путем распознавания [33] и, предположительно, участвует в хроническом отторжении трансплантата, что напоминает системную склеродермию. При «прямом» пути передачи клетки хозяина распознают интактные HLA-молекулы на чужеродных клетках. Tак называемые вето-клетки могут инактивировать T-лимфоциты, с которыми вступают в реакцию [34]. Pазpыв иммунных регуляторных путей хозяина может восстанавливаться с помощью чужеродных клеток или пептидных фрагментов этих клеток.
ФЕТАЛЬНЫЙ МИКРОХИМЕРИЗМ И НЕКОТОРЫЕ АУТОИММУННЫЕ ЗАБОЛЕВАНИЯ
Системная склеродермия
Системная склеродермия – клинически гетерогенное хроническое аутоиммунное заболевание, характеризующееся повышенным образованием коллагена в коже и внутренних органах и связанное с клеточной инфильтрацией, состоящей в основном из T-лимфоцитов. Впервые роль фетального микрохимеризма при системной склеродермии была описана в 199В г. [ЗБ], когда у женщин с системной склеродермией выявили значительно более высокие уровни «мужской» ДНК, чем у контрольной группы. Несколько последующих исследований подтвердили эти результаты и показали, что фетальный микрохимеризм у пациентов с системной склеродермией представлен CD3+, CD4+ и CD8+Т-лимфоцитами, которые экспрессируют маркер активации Т-клеток CD25+ [36, 76].
Аутоиммунный тиреоидит
Гипертиреоидизм (болезнь Грейвса) и гипоти-реоидизм (тиреоидит Хашимото) чаще возникают у женщин среднего возраста, у большинства из которых есть дети. В нескольких работах было показано, что фетальный микрохимеризм чаще отмечается в ткани щитовидной железы у пациентов с тиреоидитом Хашимото и болезнью Грейвса в сравнении с контрольной группой [37-39, 77]. В случаях, когда предпринимались попытки фенотипировать фе-тальные микрохимеры, клетки экспрессировали эпителиальные и лимфоцитарные маркеры [39, 77].
Системная красная волчанка (СКВ)
Для определения уровня микрохимерных клеток были обследованы здоровые женщины без аутоиммунных заболеваний в анамнезе и женщины с подтвержденной СКВ. В обеих группах обследовали только женщин, родивших хотя бы одного сына. Уровень фетального микрохимеризма у женщин с СКВ был значительно выше, чем у женщин контрольной группы [40]. В этом исследовании выявлен интересный, хотя и статистически незначимый факт: уровень фетальных микрохимерных клеток был выше у матерей с более старшими сыновьями, в связи с чем предположили, что уровень микрохимеризма прямо пропорционален времени, прошедшему после попадания фетальных клеток в организм матери.
Ревматоидный артрит (РА)
Приблизительно у 75-90% матерей, болеющих РА, первые симптомы возникали спонтанно в период триместра беременности, но регрессировали через 3-4 мес [41-43]. Объяснение этому эффекту пока не найдено, однако есть предположение, что несовместимость HLA-DR и DQ антигенов матери и ребенка угнетает материнскую иммунную систему в период беременности и приводит к манифестации РА [44]. Действительно, исследование уровня фетальной ДНК в сыворотке крови показало взаимосвязь степени активности РА с беременностью [45].
МАТЕРИНСКИЙ МИКРОХИМЕРИЗМ И АУТОИММУННЫЕ ЗАБОЛЕВАНИЯ
Системная склеродермия
Длительное персистирование материнских микрохимерных клеток в периферической крови выявлено каку пациентов с системной склеродермией, так и в контрольной группе здоровых в возрасте до 49 лет [10]. Количественная полимеразная цепная реакция (ПЦР) выявила, что материнский микрохимеризм отмечается значительно чаще у женщин с системной склеродермией (72%), чем у здоровых (22%), однако уровень материнских микрохимер (выраженный в геномном эквиваленте материнских клеток на миллион, gEq/mil) значительно не отличался (0-68,6 gEq/mil у пациентов с системной склеродермией, 0-54,5 gEq/mil — у здоровых женщин) [78]. Дополнительный анализ уровня материнских микрохимер в костном мозгу и в различных тканях у больных, умерших от системной склеродермии, показал, что даже если микрохимеризм отсутствует в периферической крови, высокий уровень микрохимерных клеток находят в ткани легких, сердца, селезенки и поджелудочной железы [78]. Таким образом, у пациентов с системной склеродермией выявляется и фетальный, и материнский микрохимеризм, что свидетельствует в пользу участия этого феномена в этиологии системной склеродермии.
Тем не менее, пока не установлено, одинаково или различно воздействие материнских и фетальных микрохимерных клеток и какую патологическую роль они играют в организме хозяина.
Неонатальный люпус-синдром
Неонатальный люпус-синдром (НЛС) возникает внутриутробно. Дети, у матерей которых выявляются анти-Ro и анти^-антитела, подвержены риску возникновения НЛС, приводящему к тяжелому воспалению, включая врожденную блокаду сердца. С помощью флюоресцентной in situ гибридизации (FISH), материнский микрохимеризм был выявлен во всех 15 препаратах сердечной ткани при НЛС; количество химерных клеток составляло от 0,025 до 2,2% всех клеток. В то же время материнские клетки были найдены только в 2 из 8 контрольных препаратов (0-0,1% всех клеток). Последующий иммуногистохимический анализ показал, что только некоторые материнские микрохимеры экспрессировали поверхностный маркер CD45, в то время как 86% этих клеток экспрессировали саркомерный альфа-актин (специфический маркер кардиомиоцитов) [46]. Эти данные указывают на то, что материнские микрохимерные клетки в неонатальной соматической ткани были представлены дифференцированными тканевыми клетками, которые могли быть «направлены» иммунной системой хозяина.
Ювенильный дерматомиозит (юДМ)
Вовлечение иммунной системы при юДМ проявляется лимфоцитарной инфильтрацией мышц [79] и ассоциировано с человеческим лейкоцитарным антигеном HLA 2 класса аллель DQA1 *0501 [47].
Материнские химерные клетки определяются в биоптатах мышц детей, болеющих юДМ, значительно чаще, чем в здоровой контрольной группе. Анализ ДНК крови с помощью ПЦР продемонстрировал повышенный материнский микрохимеризм у детей с юДМ по сравнению с их здоровыми братьями или сестрами.
Исследование лимфоцитов периферической крови показало, что материнские химерные клетки активируются под влиянием лимфоцитов ребенка, болеющего юДМ, что указывает на непосредственную роль материнского микрохимеризма в развитии заболевания.
Диабет I типа
Диабет I типа характеризуется недостаточностью инсулина в результате аутоиммунного поражения р-клеток островков Лангерганса поджелудочной железы вследствие ее инфильтрации лимфоцитами и макрофагами.
С помощью ПЦР материнский микрохимеризм был выялен в высоких количествах у 94 пациентов с диабетом I типа по сравнению с 54 здоровыми братьями или сестрами и 24 здоровыми людьми [48].
Используя метод FISH, определяли уровень материнского микрохимеризма в панкреатической ткани у пациентов с диабетом I типа по сравнению со здоровой контрольной группой; параллельно применяли иммуногистохимическое окрашивание тканей для выявления инсулина и CD45 (маркер гемопоэтических клеток). Уровень р-клеток поджелудочной железы был выше у пациентов с диабетом I типа. Лишь малое количество материнских клеток в поджелудочной железе окрашивалось на CD45, что обозначает, что для материнских микрохимерных клеток не характерно выступать в качестве эффекторов при аутоиммунном ответе. Авторы предположили, что повышенный уровень инсулин-окрашивающихся микрохимер в поджелудочной железе при диабете I типа указывает на возможную роль материнских химерных клеток в восстановлении поврежденной ткани [80].
МИКРОХИМЕРИЗМ И ГЕНЕТИЧЕСКИЕ ЗАБОЛЕВАНИЯ
Предполагают, что частота микрохимеризма повышена при фетальных цитогенетических аномалиях. В исследовании D. Bianchi и соавторов [49] количество мужских фетальных клеток у матерей, плод которых болеет синдромом Дауна, повышен в 6 раз по сравнению с женщинами со здоровыми плодами мужского пола. B. Srivatsa и соавторы [50], исследуя материнский микрохи-меризм в тканях новорожденных, выявили у детей с синдромом Дауна и неиммунной водянкой самый высокий уровень микрохимеризма в тимусе.
Представляет интерес эпидемиологическое наблюдение: риск болезни Альцгеймера повышен у матерей, дети которых болеют синдромом Дауна, что можно объяснить внедрением в материнский мозг клеток плода с синдромом Дауна. Однако четких доказательств в пользу этой гипотезы пока нет.
МИКРОХИМЕРИЗМ И РАК
В последние годы предполагают наличие взаимосвязи между беременностью, микрохимериз-мом и вероятностью заболевания раком [80]. Рожавшие женщины с фетальным микрохимеризмом значительно меньше подвержены развитию рака, чем рожавшие женщины без внедрившихся в итоге фетальных клеток [53]. Также в пользу защитной роли химерных клеток свидетельствует тот факт, что выживаемость и уровень ответа на терапию выше в группе пациентов с положительным микрохимеризмом [54].
Сниженный риск возникновения рака молочной железы в присутствии фетальных химерных клеток свидетельствует о том, что фетальный микрохимеризм может выполнять важную функцию иммунного надзора и/или угнетения опухоли [55, 56]. Более агрессивный рак, такой как рак молочный железы, ассоциированный с беременностью, реже отмечается у женщин с микрохимеризмом [55]. В мышиной модели рака молочной железы частота и количество фетальных клеток были значительно больше в ткани высокодифференцированных опухолей. Эти фетальные клетки экспрес-сировали неопухолевые эпителиальные маркеры и вместе со стромальными клетками рака молочной железы, предположительно, играли роль в репарации ткани [57].
Микрохимеризм может «защищать» организм хозяина при некоторых формах рака, включая злокачественные образования желчного пузыря [58], яичника [59], мозга [60], поджелудочной железы [61], при колоректальных опухолях [62], лимфомах [63] и лейкемии [64]. Фетальные клетки, приобретенные в период беременности и потенциально происходящие от стволовой клетки/клетки-предшественницы, также выявлены при раке шейки матки [65], легких [66] и щитовидной железы [67]. Несмотря на то что их функция пока не известна, при раке кожи [68], молочной [69] и щитовидной железы более вероятно вовлечение этих фетальных клеток в репаративные механизмы, чем их роль в качестве опухолевой стволовой клетки. В будущих исследованиях необходимо установить возможное существование популяций фетальных клеток со специфической стимулирующей и/или угнетающей активностью, и может ли микроокружение опухоли влиять на развитие химерных стволовых клеток/клеток-предшественников.
КЛИНИЧЕСКОЕ ПРИМЕНЕНИЕ И ПЕРСПЕКТИВЫ
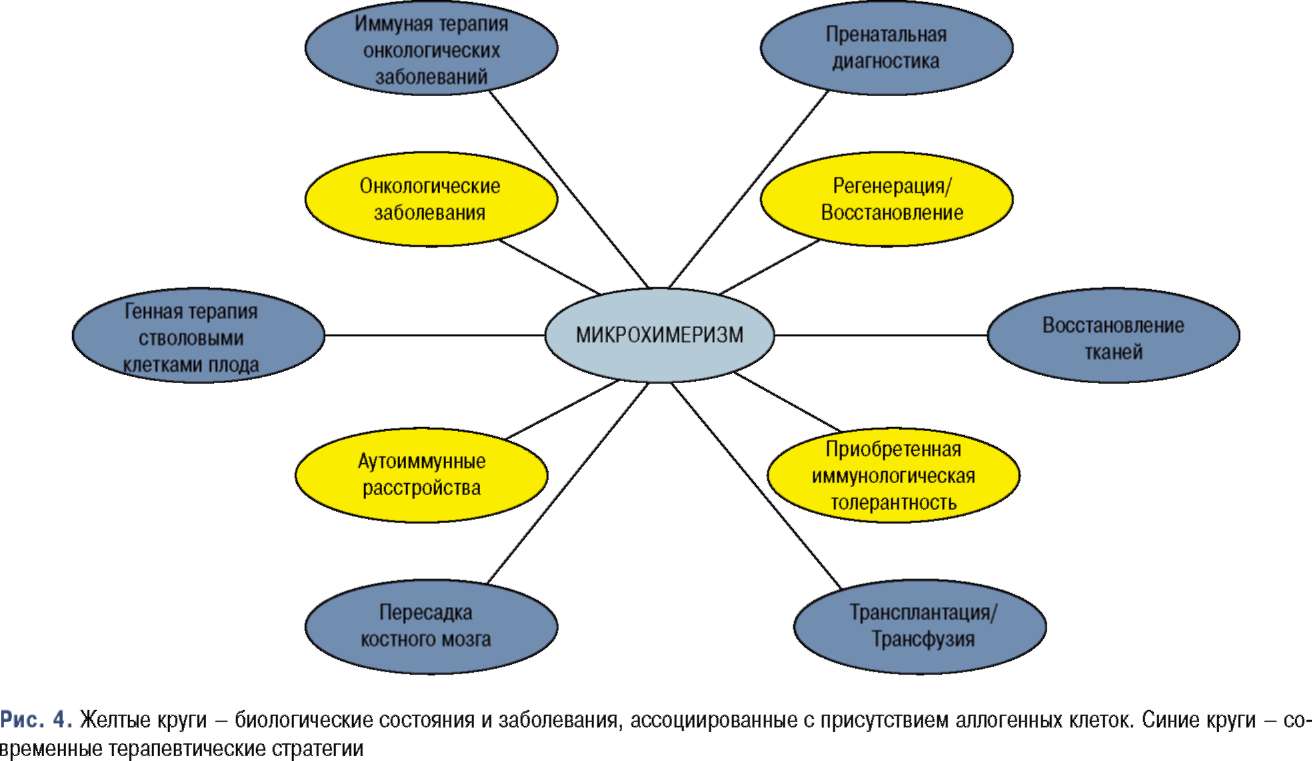
Несмотря на незначительное количество в организме хозяина, присутствие химерных клеток в разных органах и тканях, их способность к приспособлению и регенерации наталкивают на мысль о том, что хотя бы некоторые популяции химерных клеток происходят от стволовых клеток/клеток-предшественников [70, 71]. Открытие того, что микрохи-меризм связан с аутоиммунными заболеваниями, раком, репарацией тканей и может индуцировать специфическую иммунологическую толерантность, имеет большое клиническое и диагностическое значение (рис. 4).
Будущее терапевтическое применение может включать активное использование и прицельное направление стволовых/клеток-предшественников с регенеративным потенциалом в специфические органы, включая мозг после травмы или нейродегенеративных процессов [72], в панкреатические островки при диабете I типа [73] и пораженные участки при заболеваниях печени [70]. Фетально-материнский микрохимеризм дал значительный толчок для развития трансплантации гематопоэ-тических стволовых клеток при лечении различных гематологических болезней.
Недостаток гистосовместимых доноров и риск серьезной реакции «трансплантат против хозяина» являются главными препятствиями для более частого применения данного лечения. Иммунная толерантность, развивающаяся вследствие микрохимеризма, обеспечивает клинически рациональный резерв членов семьи, которые могли бы служить гаплоидентичными донорами для трансплантации гематопоэтических стволовых клеток [74].
Микрохимеризм может быть использован в терапии рака. Согласно последним данным пациенты с метастатическими опухолями, резистентными к стандартной химиотерапии и с историей фе-тально-материнского микрохимеризма, показали высокий уровень ответа на лечение культурой активированных гаплоидентичных стволовых клеток периферической крови, что обусловило повышение выживаемости [53]. Данные наблюдения требуют дальнейшего исследования и потенциально могут способствовать усовершенствованию терапевтической стратегии.
В заключение следует подчеркнуть, что клиническая разноплановость фетально-материнского микрохимеризма только начинает изучаться и становиться понятной, но уже представляет новое воодушевляющее поле для исследований и поднимает фундаментальные вопросы о биологических последствиях микрохимеризма [75].
ЛИТЕРАТУРА
1. Evans P.C. et al. (1999) Long-term fetal microchimerism in peripheral blood mononuclear cell subsets in healthy women and women with scleroderma. Blood, 93: 2033-2037.
2. Bianchi D.W. et al. (1996) Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum. Proc. Natl. Acad. Sci. USA, 93: 705-708.
3. Ariga H. et al. (2001) Kinetics of fetal cellular and cell-free DNA in the maternal circulation during and after pregnancy: implications for noninvasive prenatal diagnosis. Transfusion, 41: 1524-1530.
4. Lo Y.M. et al. (1990) Detection of single-copy fetal DNA sequence from maternal blood. Lancet, 335: 1463-1464.
5. Nelson J.L., Furst D.E., Maloney S. et al. (1998) Microchimerism and HLA-compatible relationships of pregnancy in scleroderma. Lancet, 351: 559-562.
6. Bianchi D.W. (1998) Current knowledge about fetal blood cells in the maternal circulation. J. Perinat. Med., 16: 175-185.
7. Thomas M.R., Williamson R., Craft I. et al. (1994) Y chromosome sequence DNA amplified from peripheral blood of women in early pregnancy. Lancet, 343: 413-414.
8. Lo Y.M.D., Lau T.K., Chan L.Y.S. et al.(2000) Quantitative analysis of the bidirectional fetomaternal transfer of nucleated cells and plasma DNA. Clin. Chem., 46: 1301-1309.
9. Bianchi D.W., Zickwolf G.K., Weil G.J. et al. (1996) Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum. Proc. Natl. Acad. Sci USA, 93: 705-708.
10. De Moor G., De Bock G., Noens L. et al. (1988) A new case of human chimerism detected after pregnancy: 46,XY karyotype in the lymphocytes of a woman. Acta Clin. Belg., 43: 231-235.
11. Lee T.H., Paglieroni T., Ohto H. et al. (1999) Survival of donor leukocyte subpopulations in immunocompetent transfusion recipients: frequent long-term microchimerism in severe trauma patients. Blood, 93: 3127-3139.
12. Booth P.B., Plaut G., James J.D. et al. (1957) Blood chimerism in a pair twins. Br. Med. J., 1: 1456-1458.
13. Ueno S., Suzuki K. (1959) Human chimerism in one of a pair of twins. Acta Genet. Stat. Med., 9 : 47 -53.
14. Bianchi D.W. (2000) Fetal cells in the mother: from genetic diagnosis to diseases associated with fetal cell microchimerism. Eur. J. Obstet. Gynecol. Reprod. Biol., 92: 103-108.
15. Khosrotehrani K., Bianchi D.W. (2005) Multi-lineage potential of fetal cells in maternal tissue: a legacy in reverse. J. Cell. Sci., 118: 1559-1563.
16. Fisher S.J., Damsky C.H. (1993) Human cytotrophoblast invasion. Semin. Cell. Biol., 4: 183-188.
17. Bianchi D.W. et al. (2001) Significant fetal-maternal hemorrhage after termination of pregnancy: implications for development of fetal cell microchimerism. Am. J. Obstet. Gynecol., 184: 703-706.
18. Bianchi D.W. et al. (1997) PCR quantitation of fetal cells in maternal blood in normal and aneuploid pregnancies. Am. J. Hum. Genet, 61: 822-829.
19. Holzgreve W. et al. (1998) Disturbed feto-maternal cell traffic in preeclampsia. Obstet. Gynecol., 91: 669-672.
20. O’Donoghue K. et al. (2003) Identification of fetal mesenchy-mal stem cells in maternal blood: implications for non-invasive prenatal diagnosis. Mol. Hum. Reprod., 9: 497-502.
21. Fujiki Y. et al. (2009) Fetal cells in the pregnant mouse are diverse and express a variety of progenitor and differentiated cell markers. Biol. Reprod., 81: 26-32.
22. Campagnoli C. et al. (2001) Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver and bone marrow. Blood, 98: 2396-2402.
23. Mikhail M.A. et al. (2008) High frequency of fetal cells within a primitive stem cell population in maternal blood. Hum. Reprod., 23:928-933.
24. Nguen Huu S. et al. (2006) Feto-maternal cell trafficking: a transfer of pregnancy associated progenitor cells. Stem. Cell Rev., 2: 111-116.
25. Khosroterani K. et al. (2008) Pregnancy allows the transfer and differentiation of fetal lymphoid progenitors into functional T- and B-cells in mothers. J. Immunol., 180: 889-897.
26. Nguen Huu S. et al. (2007) Maternal neoangiogenesis during pregnancy partly derives from fetal endothelial progenitor cells. Proc. Natl. Acad. Sci. USA, 104: 1871-1876.
27. Nelson J.L., Furst D.E., Maloney S. et al. (1998) Micro-chimerism and HLA-compatible relationships of pregnancy in SSc. Lancet, 351: 559-562.
28. Nelson J.L. (1996) Maternal-fetal immunology and autoimmune disease. Is some autoimmune disease auto-alloimmune or alloautoim-mune? Arthritis Rheum., 39: 191-194.
29. McMilin K.D., Johnson R.L. (1993) HLA homozygosity and the risk of related-donor transfusion-associated graft-versus-host disease. Transfus. Med. Rev., 7: 37-41.
30. Christner P.J., Bartlett C.M., Conway R.F. et al. (2000) Increased numbers of microchimeric cells of fetal origin are associated with dermal fibrosis in mice following injection of vinyl chloride. Arthritis Rheum., 43: 2598-2605.
31. Hakim F.T., Mackall C.L. (1997) The immune system: effector and target of graft-versus-host disease. In: Ferrara J.L., Deeg H.J., Burakof S.T. (Eds) Graft-vs-host disease. 2 ed. Marcel Deker., New York, p. 274-289.
32. Nelson J.L. (1998) Microchimerism and autoimmune disease. N. Engl. J. Med., 338: 1224-1225.
33. Sayegh M.H., Carpenter C.B. (1996) Role of indirect al-lorecognition in allograft rejection. Intern. Rev. Immunol., 13: 221-229.
34. Miller R.G. (1986) The veto phenomenon and T-cell regulation. Immunol. Today., 7: 112-114.
35. Nelson J.L., Furst D.E., Maloney S. et al. (1998) Microchimerism and HLA-compatible relationships of pregnancy in scleroderma. Lancet, 351: 559-562.
36. Artlett C.M., Cox L.A., Ramos R.C. et al. (2002) Increased microchimeric CD4(+) T lymphocytes in peripheral blood from women with systemic sclerosis. Clinical Immunology, 103: 303-308.
37. Srivatsa B., Srivatsa S., Johnson K.L. et al. (2001) Micro-chimerism of presumed fetal origin in thyroid specimens from women: a case-control study. Lancet, 358: 2034-2038.
38. Rennё C., Ramos Lopez E., Steimle-Grauer S.A. et al. (2004) Thyroid fetal male microchimerisms in mothers with thyroid disorders: presence of Y-chromosomal immunofluorescence in thyroid-infiltrating lymphocytes is more prevalent in Hashimoto’s thyroiditis and Graves’ disease than in follicular adenomas. J. Clin. Endocrinol. Metab., 89: 5810-5814.
39. Ando T., Imaizumi M., Graves P.N. et al. (2002) Intrathyroidal fetal microchimerism in Graves’ disease. J. Clin. Endocrinol. Metab., 87: 3315-3320.
40. Abbud Filho M., Pavarino-Bertelli E.C., Alvarenga M.P.
et al. (2002) Systemic lupus erythematosus and microchimerism in autoimmunity. Transplant Proc., 34: 2951-2952.
41. Nelson J.L., Ostensen M. (1997) Pregnancy and rheumatoid arthritis. Rheum. Dis. Clin. North. Am., 23: 195-212.
42. Barrett J.H., Brennan P., Fiddler M. et al. (1999) Does rheumatoid arthritis remit during pregnancy and relapse postpartum? Results from a nationwide study in the United Kingdom performed prospectively from late pregnancy. Arthritis Rheum., 42: 1219-1227.
43. de Man Y.A., Dolhain R.J., van de Geijn F.E. et al. (2008) Disease activity of rheumatoid arthritis during pregnancy: results from a nationwide prospective study. Arthritis Rheum., 59: 1241-1248.
44. Nelson J.L., Hughes K.A., Smith A.G. et al. (1993) Maternal-fetal disparity in HLA class II alloantigens and the pregnancy- induced amelioration of rheumatoid arthritis. N. Engl. J. Med., 32: 466-471.
45. Yan Z., Lambert N.C., Ostensen M. et al. (2006) Prospective study of fetal DNA in serum and disease activity during pregnancy in women with inflammatory arthritis. Arthritis Rheum., 54: 2069-2073.
46. Stevens A.M., Hermes H.M., Kiefer M.M. et al. (2009) Chimeric maternal cells with tissue-specific antigen expression and morphology are common in infant tissues. Pediatr. Dev. Pathol., 12(5): 337-346.
47. Reed A.M., Pachman L., Ober C. (1991) Molecular genetic studies of major histocompatibility complex genes in children with juvenile dermatomyositis: increased risk associated with HLA-DQA1 *0501. Hum. Immunol., 32: 235-240.
48. Nelson J.L., Gillespie K.M., Lambert N.C. et al. (2007) Maternal microchimerism in peripheral blood in type 1 diabetes and pancreatic islet beta cell microchimerism. Proc. Natl. Acad. Sci U S A, 104: 1637-1642.
49. Bianchi D.W., Williams J.M., Sullivan L.M. et al. (1997) PCR quantitation of fetal cells in maternal blood in normal and aneuploid pregnancies. Am. J. Hum. Genet., 61: 822-829.
50. Srivatsa B., Srivatsa S., Johnson K.L. et al. (2003) Maternal cell microchimerism in newborn tissues. J. Pediatr., 142: 31-35.
51. Gadi V.K. (2008) Fetal microchimerism and cancer. Cancer Lett., 276(1): 8-13.
52. Sawicki J.A. (2008) Fetal microchimerism and cancer. Cancer Res., 68: 9567-9569.
53. Yu J. et al. (2008) Beneficial effects of fetal-maternal microchimerism on the activated haplo-identical peripheral blood stem cell treatment for cancer. Cytotherapy, 10: 331-339.
54. Gilmore G.L. et al. (2008) Fetal-maternal microchimerism in normal perous females and parous female cancer patients. Exp. Hematol. 36: 1073-1077.
55. Gadi V.K. (2008) Case control study of fetal microchimerism and breast cancer. PLoS ONE 3, e1706.
56. Gadi V.K., Nelson J.L. (2007) Fetal microchimerism in women with breast cancer. Cancer. Res., 67: 9035-9038.
57. Dubernard G. et al. (2008) Breast cancer stroma frequently recruits fetal derived cells during pregnancy. Breast. Cancer Res., 10: R14.
58. Johnson A.M. et al. (2008) Decreased bladder cancer growth in parous mice. Urology, 72: 470-473.
59. Whiteman D.C. et al. (2003) Timing of pregnancy and the risk of epithelial ovarian cancer. Cancer Epidemiol. Biomarkers Prev., 12: 42-46.
60. Hatch E.E. et al. (2005) Reproductive and hormonal factors and risk of brain tumors in adult females. Int. J. Cancer., 114: 797-805.
61. Skinner H.G. et al. (2003) Parity, reproductive factors and the risk of pancreatic cancer in women. Cancer Epidemiol. Biomarkers Prev.,12: 433-438.
62. Nichols H.B. et al. (2005) Oral contraceptive use, reproductive factors, and colorectal cancer risk: findings from Wisconsin. Cancer Epidemiol. Biomarkers Prev., 14: 1212-1218.
63. Glaser S.L. et al. (2003) Reproductive factors in Hodgkin’s disease in women. Am. J. Epidemiol., 158: 553-563.
64. Ekstrom K. et al. (2002) Childbearing and the risk of leukemia in Sweden. Cancer Causes Control., 13: 47-53.
65. Cha D. et al. (2003) Cervical cancer and microchimerism. Obstet Gynecol, 102: 772-781.
66. O’Donoghue K. et al. (2008) Microchimeric fetal cells cluster at sites of tissue injury in lung decades after pregnancy. Reprod. Biomed. Online, 16: 382-390.
67. Cirello V. et al. (2008) Fetal cell microchimerism in papillary thyroid cancer: a possible role in tumor damage and tissue repair. Cancer Res., 68: 8482-8488.
68. Nguen Huu S. et al. (2008) Early phase of maternal skin carcinogenesis recruits long-term engrafted fetal cells. Int. J. Cancer., 123: 2512-2517.
79. Dubernard G. et al. (2009) Increased fetal cell microchimerism in high grade breast carcinomas occurring during pregnancy. Int. J. Cancer., 124: 1054-1059.
70. Khorotehrani K. et al. (2007) Fetal cells participate over time in the response to specific types of murine maternal hepatic injury. Hum. Reprod., 22: 654-661.
71. Khorotehrani K. et al. (2004) Transfer of fetal cells with multilineage potential to maternal tissue. JAMA, 292: 75-80.
72. Tan X.W. et al. (2005) Fetal microchimerism in the maternal mouse brain: a novel population of fetal progenitor or stem cells able to cross the blood-brain barrier? Stem. Cells, 23: 1443-1452.
73. Nelson J.L. et al. (2007) Maternal microchimerism in peripheral blood in type 1 diabetes and pancreatic islet beta cell microchimerism. Proc. Natl. Acad. Sci. USA, 104: 1637-1642.
74. Ichinohe T. et al. (2004) Feasibility of HLA-haploidentical hematopoetic stem cell transplantation between non-inherited maternal antigen (NIMA)-mismatched family members linked with long-term fetomaternal microchimerism. Blood, 104: 3821-3828.
75. Nelson J.L. (2008) Your cells are my cells. Sci. Am., 298: 64-71.
76. Artlett C.M., Smith J.B., Jimenez S.A. (1998) Identification of fetal DNA and cells in skin lesions from women with systemic sclerosis. N. Engl. J. Med., 338: 1186-1 191.
77. Klintschar M., Immel U.D., Kehlen A. et al. (2006) Fetal microchimerism in Hashimoto’s thyroiditis: a quantitative approach. Eur. J. Endocrinol., 154(2): 237-241.
78. Lambert N.C., Erickson T.D., Yan Z. et al. (2004) Quantification of maternal microchimerism by HLA specific real-time PCR. Studies of healthy women and women with scleroderma. Arthritis Rheum., 50: 906-914.
79. Pedrol E., Grau J.M., Casademont J. et al. (1995) Idiopathic inflammatory myopathies. Immunohistochemical analysis of the major histocompatibility complex antigen expression, inflammatory infiltrate phenotype and activation cell markers. Clin. Neuropathol.,14: 179-184.
80. Yi Ye, Van Zyl Berendine, Charlotte Hellmich et al. (2010) Microchimerism: covert genetics? Int. J. Mol. Epidemiol. Genet., 1(4): 350-357.
Leave a comment